




第2回 理論のなかのシステム
-

徳安 彰AKIRA TOKUYASU
法政大学社会学部教授東京大学文学部社会学科卒業。同大学院社会学研究科博士課程修了。法政大学社会学部専任講師、同助教授、ビーレフェルト大学客員研究員、法政大学社会学部教授、法政大学社会学部学部長を歴任。共著に『社会理論の再興』(ミネルヴァ書房)、『理論社会学の可能性』(新曜社)など。
第2回は、システム理論やサイバネティクスにおけるシステムの考え方を、いくつかの代表的な理論について素描する。それらの理論は、社会システム理論の展開とも密接な関係をもっている。また、もともとの理論はおおむね生物と機械の原理を抽象的に記述するものであって、思想的、イデオロギー的含意はないが、それが社会システム理論にもち込まれると、一定の思想的、イデオロギー的含意をもつようになる。その点にも触れながら、ホメオスタシスを出発点にしてネガティブ・フィードバックとポジティブ・フィードバックの対概念について検討し、つぎに最小多様度の法則をシステム/環境−関係と関連させながら考察し、その延長線上にオートポイエーシスを位置づける。それによって、ルーマンの社会システム理論を理解するための理論的文脈が明らかになるだろう。
はじめに

第1回の「思想のなかのシステム」では、システムは要素の総和とは異なるものであるという全体論的な考え方を基礎にして、社会システムがたんに要素となる人間(あるいは人間の行為、コミュニケーション)とは異なるだけでなく、人間を抑圧し、疎外するものになりうるという考え方を、社会思想や古典社会学を中心に見てきた。またこの抑圧や疎外は、社会システムが合理的に作られているにもかかわらず、ではなく、合理的に作られているからこそ生じる、ということも見てきた。
第2回の「理論のなかのシステム」では、システム理論やサイバネティクスのなかに見られるシステムの考え方を見ていく。システム理論やサイバネティクスは、自然科学や工学の研究のなかから生まれてきた理論である。研究の主要な対象は、自然物や自然現象、あるいは人工物(おもに機械)であり、そこで得られた知見を一般化することによって、人間や社会をも対象とする理論を構築しようとしてきた。
システム理論やサイバネティクスでも、合理性が問題となる。この場合の合理性の第一の意味は、研究対象を論理的に説明することができる、という科学的認識のあり方にかかわる。およそ科学は、対象を説明する理論が論理的一貫性をもたなければならないからである。また、科学理論が対象を合理的に説明することができるならば、そのかぎりにおいて、対象そのものが合理的に作動している、と考えることができる。自然物や自然現象の科学的探求としての自然科学は、このような観点のもとで発展してきた。
他方、工学は人工物を構築することにかかわる。この場合、合理性とは、人工物が合理的に作動する、ということであると同時に、人工物が合理的に作動するように設計、制御することを意味する。自然物や自然現象を対象とする自然科学が、いわば自然のなかの合理的な設計図を読み取ろうとしているのに対して、工学は人間自身が合理的な設計図を書いて、そのとおりに人工物を組み立て、合理的に動くように制御しようとする。なぜなら、一般に人工物は、人間の生活に役立つという目的のために作られるのであり、そのかぎりにおいて、目的に適合するという意味で合理的に作動しなければならないからである。
自然物や自然現象についても、その因果的なメカニズムが解明されてくると、そこに介入して、人間の生活に役立つように制御しようという工学的な発想が生まれてくる。たとえば、経験的な知識にもとづいて交配によって新しい品種を開発する古典的方法から、遺伝子配列を直接操作する現代の遺伝子工学にいたるまで、生命科学では工学的な方法が取り入れられている。人間や社会についても、行動主義心理学は、人間の行動の因果的なメカニズムが解明されれば、人間を意のままに成育させたり、思うように行動を操ったりすることができるはずだと考えたし、社会工学は、社会現象の因果的なメカニズムが解明されれば、社会を思うような方向に発展させることができるはずだと考えた。
こうした自然科学や工学を基礎にしたシステム理論やサイバネティクスにとって、いかなるものであれ、システムは最初から人間に外在するものであり、その意味で最初から人間にとっては異物であった。しかしシステムは、社会思想や古典社会学のいう意味で人間を抑圧したり疎外したりする存在ではなく、むしろ人間が制御する可能性をもった対象だった。人間が考えた目的に適合するようにシステムを設計することができるかぎりにおいて、人間は主体性を発揮することができるし、システムを制御することができるかぎりにおいて、やはり人間が主体となることができる。
それでは、システム理論やサイバネティクスの知見を導入したルーマンの社会システム理論は、社会工学と同じように社会を技術的に制御しようとする理論なのだろうか。ハーバーマス−ルーマン論争を収めた本の原題は、Theorie der Gesellschaft oder Sozialtechnologie – Was leistet die Systemforschung?(社会の理論か社会工学か──システム研究は何をするのか?)となっており、ルーマンの社会システム理論が工学的なものであるかどうかが争点になっていることを示唆している(Habermas/Luhmann, 1971=1987)。この問題については、後の回でルーマンの理論を論じるときに、あらためて詳しく検討することにしよう。
だが、そもそもルーマンは、導入したシステム理論やサイバネティクスの特定の理論をそのまま社会システムに適用するのではなく、独自の一般化や読み替えをした上で適用している。じつに一筋縄ではいかない。だから、ルーマンの理論を理解するために、熱心にもとのシステム理論やサイバネティクスを理解しても、かえって違和感さえ覚えてしまう。それは、ルーマン以前の社会システム理論の代表者であるパーソンズも同じである。とはいえ、いわゆる元ネタを無視しては、彼らの理論を十分に理解することはできない。そこで、まずはパーソンズが参照したシステム理論やサイバネティクスの理論から出発して、ルーマンが初期に言及したアシュビーの最小多様度の概念、後期のルーマンの代名詞となったオートポイエーシスの理論を中心に見ていくことにしよう。
ホメオスタシスとネガティブ・フィードバック
ルーマン以前の社会システム理論の代表者はタルコット・パーソンズ(Talcott Parsons, 1902-1979)である。パーソンズが当時のシステム理論からとり入れた第一の概念は、生理学者のキャノン(Walter Bradford Cannon, 1871-1945)が提唱したホメオスタシスだった。ホメオスタシスは、恒常性あるいは恒常性維持と訳される。キャノンは、生物有機体にさまざまな恒常性維持の機能が備わっていることを体系的に理論化した。(本節におけるホメオスタシスと社会システム理論の関係についての概説は、[徳安, 2017]を参照)
ホメオスタシス概念の有力なルーツは、19世紀フランスの生理学者ベルナール(Claude Bernard, 1813-1878)の内部環境論である。ベルナールは、生物の体内にあって組織細胞や器官を包む血液やリンパ液のような体液を、細胞にとっての内部環境と名づけ、内部環境の固定性ないし安定性が生命維持の条件であると考えた。1920年代に、キャノンはベルナールの考え方を生命維持の基本的原理に展開し、これをホメオスタシス(homeostasis)と名づけた。キャノンは、著書『からだの知恵』(Cannon, 1932=1981)のなかで、血液中の水の量、血液中の塩分の量、血液中の糖、血液中のタンパク質、血液中の脂肪、血液中のカルシウム、十分な酸素の量、血液の中性、体温といった要素が恒常的に維持されるメカニズムを示し、これらが全体として生物有機体の生命を維持する働きをしていると述べた。
たとえば、血液中の糖(血糖値)の恒常性は、図1のようなメカニズムで維持されている。血糖値は高すぎても低すぎても生命維持に支障をきたすが、じっさいには食事の仕方などによって上昇したり低下したりする。血糖値を一定の範囲に維持するために、上昇すればそれを抑制するメカニズムが働き、低下すればそれを抑制する別のメカニズムが働く。([Cannon, 1932=1981, 訳pp.109-133]を参照。ただし、図はかならずしもキャノンの記述通りではない)
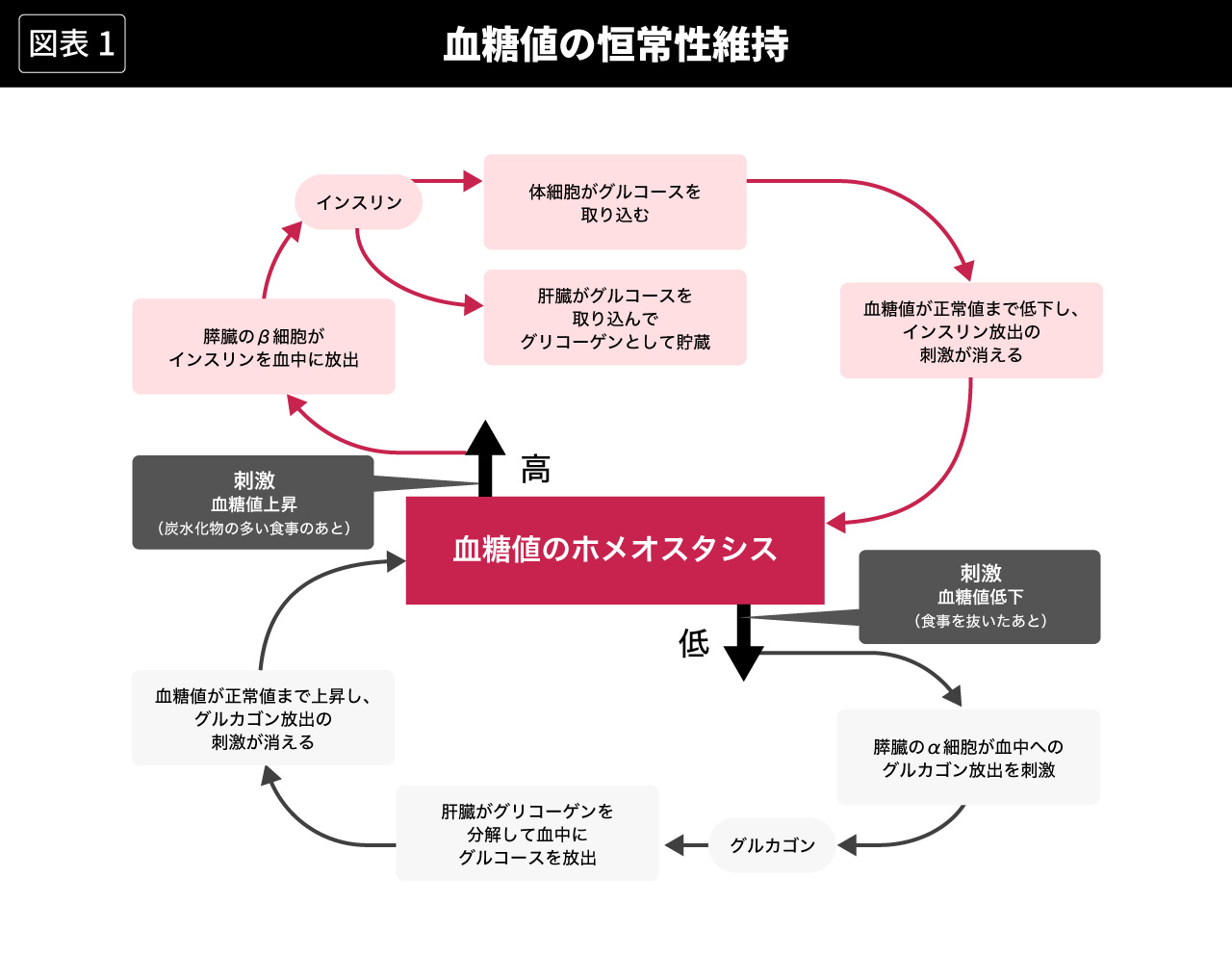
生物有機体の安定した状態を確保するためのさまざまな生理学的作用について、キャノンは要約的に次のように述べている。
生体のなかで、安定した状態の主要な部分を保つ働きをしている、相互に関連した生理学的な作用は、ひじょうに複雑であり、また独特なものなので──それらのなかには、脳とか神経とか心臓、肺、腎臓、脾臓が含まれ、すべてが協同してその作用を営んでいる──私はこのような状態に対して恒常状態(ホメオステーシス(homeostasis))という特別の用語を用いることを提案してきた。この用語は、固定し動かないもの、停滞した状態を意味するものではない。それは、ある状態──変化はするが相対的に定常的な状態──を意味するものである。(Cannon, 1932=1981, 訳pp.27-28)
システム理論の用語でいいかえれば、生物有機体は、外部環境と物質やエネルギーの交換(代謝)を行う開放システムであり、絶えず環境からの影響(攪乱)をうけながら、自己の定常状態を維持するためのフィードバック・メカニズムをもっている、ということができるだろう。またその定常状態は、まったく固定した静的平衡ではなく、一定の許容範囲内で変動する動的平衡であり、そのかぎりにおいてシステムは安定的とみなすことができる。さらに、発達した複雑なシステムとしての生物有機体は、複数のフィードバック・メカニズムをもっており、それらが協働することによって、全体としての恒常性を保っている。
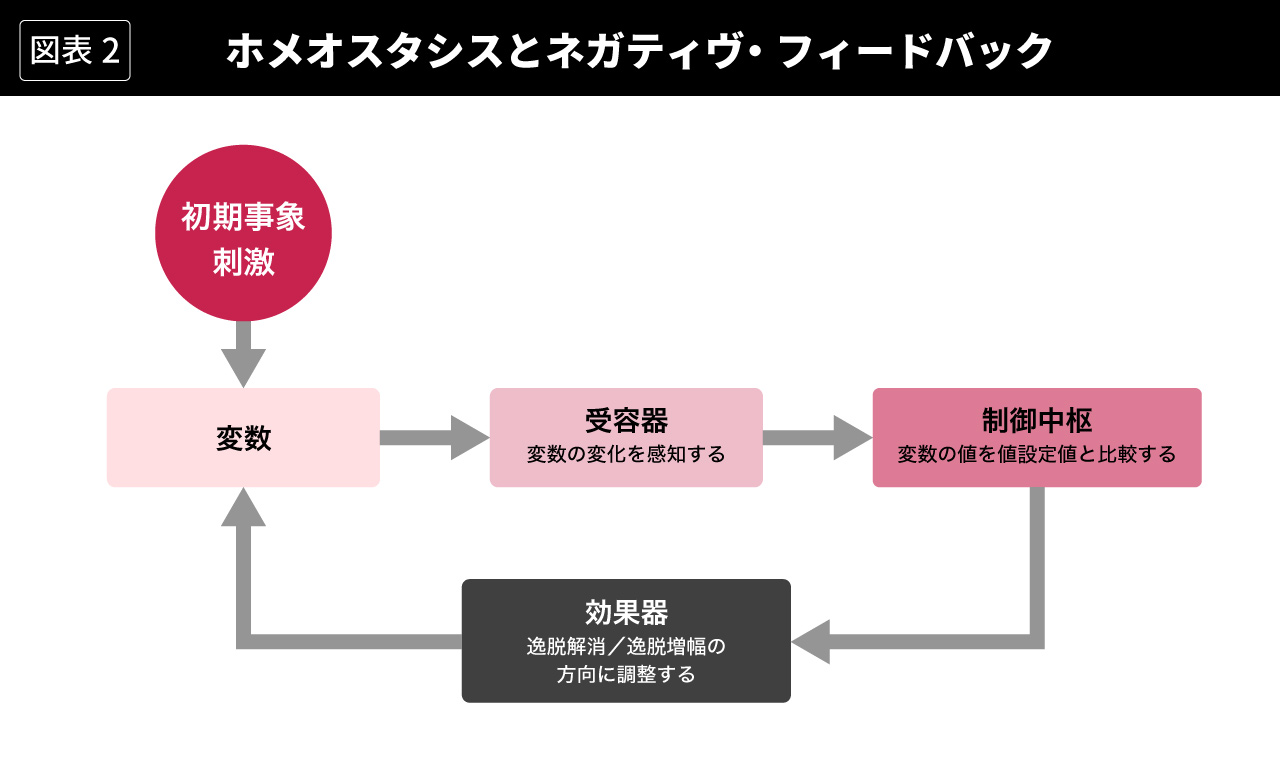
これを受けて、20世紀半ばに、N.ウィーナーやW. R.アシュビーがサイバネティクスの観点から、L. v.ベルタランフィが一般システム理論の観点から、ネガティブ・フィードバック(負のフィードバック)による制御メカニズムを備えたシステムの自己維持メカニズムとして、ホメオスタシスを定式化した(Wiener, 1961; Ashby, 1961; Bertalanffy, 1968)。このメカニズムを図式化すると、図2のように表現することができる。生物有機体でいえば、図2の「変数」が図1の血糖値にあたる。初期事象の刺激によってこの変数の値が変化すると、それを「受容器」が関知して「制御中枢」に伝達し、「制御中枢」は変数の値を設定値(生物有機体では生命維持のために望ましい値)と比較して「効果器」に調整の指令を出し、「効果器」は逸脱解消の調整をする、というわけである。
ここで重要なのは、このフィードバック・メカニズムが、初期事象の刺激による変数の変化を打ち消す方向に働き(それが「ネガティブ」という形容詞の意味である)、それによってシステムの恒常性を維持するという点である。つまり、このフィードバック・メカニズムは、システムの変数が設定値から大きくはずれないように(逸脱を解消するように)制御することによって、システムの状態を安定的に維持するのである。キャノンのホメオスタシスでは、自律神経系と内分泌系の制御によって血液の成分、体温などが一定範囲内に維持されている状態がそれである。サイバネティクスでは、たとえばサーモスタットのついた空調機によって室温が一定範囲内に維持されている状態がそれである。
このようにホメオスタシスとネガティブ・フィードバックを明確に結びつける考え方は、次のウィーナーの文章のなかに明確に述べられている。
われわれが生きつづけることができるのは、ひじょうに特殊な環境の中においてであり、その環境をわれわれは身のまわりにつけてゆくが、やがてわれわれのからだは自己を再構成してゆける限度より速い速度で崩壊し始める。そしてわれわれは死ぬ。もし体温が正常値である華氏九八・六度より一度も上昇するか下降すれば、われわれはそれに気づきはじめ、もし一〇度も高くなるか低くなれば、われわれは確実に死ぬ。われわれの血液中の酸素や炭酸ガスや内分泌腺から流れでるホルモンはすべて、その水準に不都合な変化が起こることに抵抗する機構によって制御されている。これらの機構は、ホメオスタシスと呼ばれるものを構成しており、いずれも、自動機械で実現できる型の負のフィードバック機構である。(Wiener, 1950=2007, 訳pp.99-100)
ここまでくれば、社会の秩序を維持するメカニズムも、このホメオスタシスとネガティブ・フィードバックによって理論的に表現することができるのではないかという発想は、きわめて自然なことに思われるだろう。じっさい、キャノン自身が『からだの知恵』のエピローグで、社会的恒常性について語っている(Cannon, 1932=1981, 訳pp.322-344)。こうしたホメオスタシスとネガティブ・フィードバックの概念からヒントを得て、パーソンズは、社会システムが環境の変動に対して内部のパターンの恒常性を維持すると考えた。
パーソンズの場合、社会システムの状態を安定的に保つネガティブ・フィードバックのメカニズムは、社会統制と呼ばれる。社会統制とはsocial controlの訳であり、システム理論であれば「社会制御」と訳すことができるし、その方がサイバネティクスとの関連をより明確に表現できるのだが、ここでは社会学の慣習的訳語にしたがっておく。社会統制は、逸脱に対して発動されるフィードバック・メカニズムである。
逸脱と社会統制のメカニズムとは、その準拠点として、個人行為者を考えるか、それとも相互行為システムを考えるかにしたがって、二つの仕方で定義できる。第一の文脈では、逸脱とは、行為者が一つまたはそれ以上の制度化された規範的パターンに違反して、行動するように動機づけられる傾向であり、これにたいして社会統制のメカニズムとは、この行為者の行動やかれが相互行為している他者の行動の動機づけられる過程で、こうした逸脱への傾向を妨げることになる。第二の文脈つまり相互行為システムの文脈では、逸脱とは(静的均衡であれ動的均衡であれ)、相互行為過程の均衡を攪乱するようなぐあいに、そのシステムの構成員たる行為者の一人またはそれ以上の人びとが行動する傾向である。したがってその逸脱は、相互行為システムの状態の変動か、または対抗勢力による再均衡のいずれかに帰着する傾向によって定義される。後者が社会統制のメカニズムなのである。ここではこうした均衡は、つねに、多かれ少なかれ制度化されている規範的パターンの体系と行為との統合を意味していると仮定される。(Parsons, 1951=1974, 訳p.252:一部改訳)
この文章から明らかなことは、社会統制はネガティブ・フィードバックによって逸脱を妨げ、システムを均衡(平衡)状態に戻そうとするメカニズムだということである。ただし、パーソンズの場合には、行為者の行為のレベルと相互行為システム(=社会システム)の状態の二つのレベルで社会統制が考えられており、さらに逸脱が規範的パターンからの逸脱として概念化されている点に注意が必要である。この規範的パターンは、生物有機体や機械では出てこない要素であり、社会学の独自性を意味すると同時に、規範的パターンに同調する方が逸脱するよりも望ましいという含意を生むことになった。
キャノンのホメオスタシスにおいて、生物有機体にとっては恒常性を維持することが生存にとって必要であるように、パーソンズの社会システム理論においても、社会システムにとっては均衡(平衡)を維持することが存続にとって必要であると考えられている。そうすると、特定の規範的パターンを所与とした場合、この均衡(平衡)維持は現状維持と同じ意味になり、革新や発展のような事態も闘争や破壊のような事態も、ともに規範的に望ましくない逸脱とみなされて統制されることになる。この点が、1950年代から1960年代にかけての社会の既存秩序に対する疑問、批判、反抗の機運の高まりのなかで、パーソンズの社会システム理論に保守主義や体制派というレッテルを貼って、イデオロギー的に批判する動きにつながった(富永, 2008, pp.499-501)。
だがこのコラムでは、パーソンズの社会システム理論にあまり深入りすることはできない。ここでは、パーソンズが、ホメオスタシスとネガティブ・フィードバックの概念をとり入れて、社会システムの安定的な均衡(平衡)状態を重視する理論を展開したことを確認するにとどめておこう。
ポジティブ・フィードバックとモルフォジェネシス
ネガティブ・フィードバックのメカニズムとは対照的なものとして考え出されたのが、ポジティブ・フィードバックのメカニズムである。ポジティブ・フィードバックのメカニズムも、先の図2で表すことができる。ネガティブ・フィードバックでは、初期事象の刺激によって変数の値が変化すると、それを「受容器」が関知して「制御中枢」に伝達し、「制御中枢」は変数の値を設定値と比較して「効果器」に調整の指令を出し、「効果器」は逸脱解消の調整をするという過程をたどるが、ポジティブ・フィードバックでは、制御中枢が効果器に出す調整の指令が逆向きになり、「効果器」が逸脱増幅の調整をする。
実用的な工学の分野では、ネガティブ・フィードバックのメカニズムが、すでに18世紀末に蒸気機関を発明したワット(James Watt, 1736-1819)の遠心調速機のようなかたちで実装されていたように、ポジティブ・フィードバックのメカニズムも、すくなくとも20世紀初頭にはラジオの再生回路における信号増幅メカニズムとして実用化されていた。また、生物にもこのポジティブ・フィードバックのメカニズムが備わっており、たとえば妊婦の出産の時期が迫ると、胎児が子宮頸部を圧迫し、子宮頸部から脳に刺激が伝達されると、脳下垂体後葉からオキシトシンが分泌され、オキシトシンによって子宮の収縮が促進されるというサイクルのくり返しによって、最終的に胎児が子宮から押し出されて出産にいたる。
このポジティブ・フィードバックのメカニズムを、サイバネティクスの原理として明確に提示したのが、マゴロウ・マルヤマ(丸山孫郎)である。
サイバネティクスはその成立時以来、多かれ少なかれ自己調整的、平衡化的なシステムの科学と見なされてきた。恒温器、体温の生理学的調整、自動操縦装置、経済的・政治的諸過程が、逸脱解消的(deviation counteracting)フィードバック・ネットワークの数学的モデルに基づいて研究された。しかしサイバネティクス研究家たちは、相互的因果関係の逸脱解消的な側面に焦点を定めることにより、相互因果の効果が逸脱増幅的(deviation amplifying)であるようなシステムにはあまり関心を寄せなかった。しかしこの逸脱増幅を示すシステムはいたるところに見られる。生命有機体の進化、様々なタイプの文化の勃興、情緒障害を生む対人関係、国家間の紛争、そして「悪循環」や「複利」などと漠然と呼ばれている過程、あるいは偶然な発端の始動(initial kick)を増幅させ、逸脱をつみ重ね、当初の状態から遠ざかる相互的因果関係の過程である。(Maruyama, 1963=1987, 訳pp.81-82)
マルヤマは、もっぱらネガティブ・フィードバックのメカニズムに依拠した研究をファースト・サイバネティクスと呼び、ポジティブ・フィードバックのメカニズムに依拠した研究をセカンド・サイバネティクスと呼んだ。また、ネガティブ・フィードバックによる逸脱解消的な因果関係を「形態維持(モルフォスタシス(morphostasis))」と呼び、ポジティブ・フィードバックによる逸脱増幅的な因果関係を「形態形成(モルフォジェネシス(morphogenesis))」と呼んだ(Maruyama, 1963=1987, 訳p.82)。
ところで、システムの恒常性維持と変化はどちらもありうる現象である。生物有機体で恒常性が失われると、たいていは衰弱か死にいたる。低体温症や高熱の状態を思い浮かべてみればわかる。機械も、たとえば際限なく室温を上げたり下げたりするのは、サーモスタットの故障であり、部屋にいる人間にとってははなはだ都合が悪い。ホメオスタシスとネガティブ・フィードバックは、自然物についてはそれじたいにとって(とくに生物有機体にとって)、人工物ではそれを利用する人間にとって、きわめて重要なメカニズムである。その意味できわめて合理的である。じっさい、社会システムについても、革新や発展はシステムの新しい状態をもたらすかもしれないが、それが望ましい状態かどうかは一概にいえないし、闘争や破壊は現状を変えるとしても、往々にして無秩序をもたらして、システムの崩壊を招きかねない。
逸脱増幅の結果の成否を決めるために必要なのが、超安定性(ultrastability)という考え方であり、それを示したのが図3である。ネガティブ・フィードバックによって逸脱解消がうまく行われれば、システムの変数は一定の設定値の範囲内で平衡(均衡)状態1を保ち、そのかぎりにおいてシステムは安定的である。しかし、ポジティブ・フィードバックによって逸脱増幅が起こると、システムの変数は既存の設定値の範囲内にとどまることができなくなる。この場合に、変数の設定値が変更されて、異なる設定値の範囲内で平衡(均衡)状態2が保たれるようになれば、システムは超安定的といえる。しかし、そのためには新しい変数の許容範囲を設定するメカニズムがシステムに内蔵されていなければならず、さもないとシステムは崩壊または死にいたってしまう。(超安定性の条件のより厳密な説明については、[Ashby, 1960=1967, 訳pp.95-160]を参照)
ポジティブ・フィードバックのメカニズムによるシステムの変化、成長、発展、進化といった現象についての説明が、サイバネティクスに新しい可能性を開いたことは疑いない。しかし、ポジティブ・フィードバックがつねにシステムのよりよい状態をもたらしてくれるとはかぎらず、場合によってはシステムの崩壊や死をもたらすことに留意しなければならない。とくに、この概念が社会システムに適用される場合、変化、成長、発展、進化が無条件に肯定的に評価され、ある平衡状態に安定的にとどまることが保守、停滞として否定的に評価される傾向がある。パーソンズのいう「慣性の法則」などもってのほかだというわけである。だが厳密に考えれば、生物有機体がそうであるように、社会システムの働きもどちらか一方のフィードバック・メカニズムだけで説明し尽くすことはできない。複雑な社会システムは、複雑な生命有機体と同じように両方のフィードバック・メカニズムをもっていると考えるべきだろう。

すこし脇道にそれるが、二つのフィードバックの概念は、マネジメントの分野でいささか異なる使い方がされている。語感だけでいえば、ポジティブ・フィードバックは肯定的なフィードバックのようにも思われるせいか、従業員の仕事の良い部分を肯定的な言葉で評価することによってモチベーションを上げる、という意味で用いられる。ぎゃくにネガティブ・フィードバックは否定的なフィードバックのように思われるせいか、従業員の仕事の悪い部分を否定的な言葉で評価することによってモチベーションを下げる、という意味で用いられる。これが、上記で説明したようなサイバネティクスの本来の意味でのフィードバックの2類型と異なることは明らかだろう。(たとえば[グロービス経営大学院 MBA用語集]を参照)
最小多様度とシステム/環境−関係
W. R.アシュビーも、サイバネティクスの観点から、生物と機械に共通する原理をホメオスタシスに見た。つまり、基本的にはネガティブ・フィードバックのメカニズムを備えて、自己を維持するシステムを考えた。その意味では、アシュビーの理論についてあらためて冗長な説明をくり返す必要はない。だが、アシュビーは、ホメオスタシスの説明に多様度(variety)という概念を持ち込んだ点がユニークであり、これがルーマンの社会システム理論における複雑性の概念につながっていく。その点に留意しながら、アシュビーの考え方を見ていこう。
多様度の概念は、さまざまな可能性の集合という考え方にもとづいて、その集合の要素の数として定義される。多様度を数え上げるのは簡単なことに思われるかもしれないが、多様度はその集合に内在する特性ではなく、観察者とその識別能力が指定されなければならないことに、アシュビーは注意を促している(Ashby, 1956=1967, 訳p.154)。つまり、多様度は世界の存在や現象の客観的な性質ではなく、観察によってはじめて定義される、観察者にとっての主観的な世界のありようなのである。アシュビーは、この点をかならずしも認識論的な議論として展開していないが、ルーマンの社会システム理論ではこの点が重要になってくる。
つぎに、二つの集合間の関係で、一つの条件下にある多様度が、他の条件下にある多様度よりも少ないときに生じるものとして、拘束(constraint)の概念が定義される。具体例として、人間の性の多様度に対する男子校の性の多様度の少なさ(というより多様度ゼロ)、イギリスの交通信号燈の3色の点灯・消灯のじっさいの組合せが、3色が独立に点灯・消灯する場合のすべての組合せ(8通り)よりも少なく、4通りしかないことが、挙げられている(Ashby, 1956=1967, 訳p.157)。さらに、拘束があるときにはいつでも利得がえられるとして、科学におけるすべての自然法則はひとつの拘束であると述べている。
科学は法則を求める。それゆえに科学は拘束を探し出すことに関心をもつ。(ここでより大きな集合とは、その行動が自由で無秩序ならば、生じたかもしれないものから成り、より小さな集合とは実際におこるものから成っている)。‥‥サイバネティクスは全体をあらゆる可能な豊かさで考察し、そして現実がなぜ全体の可能性の或る部分に制限されなければならないかを尋ねるのである。(Ashby, 1956=1967, 訳pp.161-162)
先回りしていえば、この拘束の概念がルーマンにおける複雑性の縮減という考え方につながっていくことは、比較的わかりやすいだろう。では、この多様性と拘束の概念は、ホメオスタシスの考え方とどのように関係してくるのだろうか。アシュビーは、生物の生き残りとは、生きている生物に対応する状態M1, ‥‥Mkが、ある本質的変数(essential variable)が指定された(生理学的な)限界内に保たれているような状態であるとしている(Ashby, 1956=1967, 訳p.243)。本質的変数の例として血液中の糖、塩分、タンバク質、脂肪、カルシウムの量や濃度を思い浮かべれば、この考え方がキャノンのホメオスタシスと同じであることは明らかであろう。アシュビーはそのような働きを調整と呼び、「一般にすぐれた調整装置の本質的な特徴は攪乱因から本質的変数へと多様度が流れることを妨げることである」と述べている(Ashby, 1956=1967, 訳p.248)。
ここで、攪乱因のとりうる手の集合をD、調整装置のとりうる手の集合をRとすると、結果の集合の多様度は、〈Dの多様度/Rの多様度〉で表すことができる。Rの打つ手が変わらなければ、結果の多様度はDの手の多様度に等しくなる。Rの打つ手が2つになれば、結果の多様度は半分になり、Rの打つ手が3つになれば、結果の多様度は3分の1になる。以下同様で、結果の多様度をある数にまで減少させるためには、Rの多様度がある適当な最小値以上でなければならない。これが最小多様度の法則(Law of Requisite Variety)の法則である。「Rの多様度だけがDに由来する多様度をひき下げることができる。多様度だけが多様度を破壊することができる」というわけである(Ashby, 1956=1967, 訳pp.255-256)。
もう一度、この最小多様度の法則をホメオスタシスとつなげて考えると、ある本質的変数が一定の状態の多様度の範囲内に保たれるためには、攪乱因の多様度を本質的変数の多様度にまでひき下げることのできる多様度を持った調整装置が必要だ、ということになる。攪乱因が環境に由来すると考え、攪乱因の多様度を適切に調整する(つまり多様度をひき下げる)調整装置と、その結果として一定に保たれる本質的変数をシステム内部の構成要素と考えると、最小多様度とホメオスタシスは図4のように表すことができる。
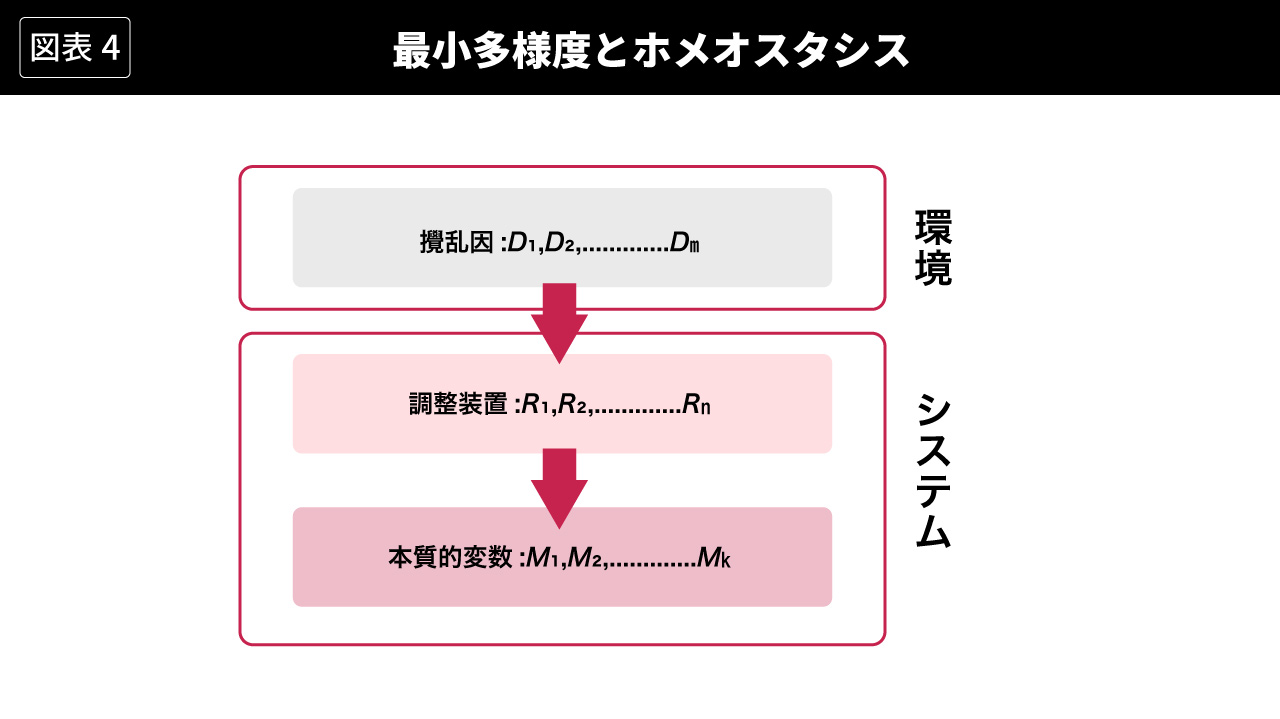
最初に多様度の定義を示したときに、アシュビーが多様度を、世界の客観的な性質ではなく、観察者の主観的な認知とみなしたことを指摘した。しかし、アシュビー自身は、多様度は観察者である研究者が操作的に定義するものとみなしており、想定されるシステム自体が環境について認知するものとはみなしていない。図4における多様度も、研究者が操作的に定義したものとして示されている。この操作的な定義にもとづいて、たとえばmとnとkの大小関係を数学的に定式化することができるわけであり、システムが存続する(生物ならば生き残る、つまりホメオスタシスを実現する)ためのkの値が与えられれば、nの最小値(つまり最小多様度)を求めることができることになる。
この図式の認知的な部分を、研究者から見たシステム/環境−関係ではなく、システムから見たシステム/環境−関係に読み替えた例として、ユクスキュル(Jakob von Uexküll, 1864-1944)の理論が挙げられる。ユクスキュルは、共著書『生物から見た世界』(Uexküll/Kriszat, 1934=2005)の冒頭で、マダニの認知と行動の例を挙げて、つぎのように述べている。
雌は交尾を終えると、八本肢を総動員して適当な灌木の枝先までよじのぼる。これは、十分な高さから下を通りかかる小哺乳類の上に落ちるか、大型動物にこすりとられるかするためである。
この目のない動物は、表皮全体に分布する光覚を使ってその見張りやぐらへの道を見つける。この盲目で耳の聞こえない追いはぎは、嗅覚によって獲物の接近を知る。哺乳類の皮膚腺から漂い出る酪酸の匂いが、このダニにとっては見張り場から離れてそちらへ身を投げろという信号として働く。そこでダニは、鋭敏な温度感覚が教えてくれるなにか温かいものの上に落ちる。するとそこは獲物である温血動物の上で、あとは触覚によってなるべく毛のない場所を見つけ、獲物の皮膚組織に頭から食い込めばいい。こうしてダニは温かな血液をゆっくりと自分の体内に送りこむ。
‥‥
このダニにとってたっぷりの血のごちそうはまた最後の晩餐でもある。というのは、彼女にはもう、地面に落ちて産卵し死ぬほかになにもすることがないからだ。(Uexküll/Kriszat, 1934=2005、訳pp.12-13)
この例では、生物個体のホメオスタシスではなく、種の保存のための行動が問題になっている。この例について、ユクスキュルは、ダニの行動をたんなる機械のように客観的に理解するのではなく、むしろダニを主体として客体である哺乳動物との関係を見なければならないと述べている。もちろん、ダニの行動は生物としての客観的な構造によって規定されているのだが、ここで問題となるのは、主体としてのダニがみずからの行動のために周囲の世界をどのように捉えているか、ということである。つまり、研究者から見た客観的なダニの周囲の世界のありようが問題なのではなく、ダニから見た「主観的な」周囲の世界のありようが問題なのである。すこし文学的にいえば、ダニによって生きられる世界が焦点である。
ダニを取り囲む豊かな世界は崩れさり、重要なものとしてはわずか三つの知覚標識と三つの作用標識からなる貧弱な姿に、つまりダニの環世界に変わる。だが環世界のこの貧弱さはまさに行動の確実さの前提であり、確実さは豊かさより重要なのである。(Uexküll/Kriszat, 1934=2005、訳p.22)
この引用文でいう「環世界」はドイツ語のUmweltの訳であり、通常は英語のenvironmentと同義とみなされて「環境」と訳されることが多い。だがユクスキュルの場合には、客観的な環境にはUmgebungという語があてられ、それと区別される生物に認知される主観的な環境をさす語としてUmweltがあてられている。つまり、当該生物が認知する、みずからを取り巻く世界がUmweltなのである。アシュビーの多様度の概念をもちいれば、生物が主観的な認知によって客観的な環境の多様度をみずからにとっての環世界の多様度にひき下げ、それに対応した行動の多様度によって種の保存を確保している、といえるだろう。このような「主観的」多様度こそ、ルーマンの社会システム理論における複雑性の概念につながるものである。
オートポイエーシスと認識
ここまでの議論で暗黙の前提になっていたのは、環境のなかのシステムだった。つまり、システムは環境からの影響をうけながら、ネガティブ・フィードバックによって自己の状態を一定に維持したり、ポジティブ・フィードバックによって状態を変えたりするという意味で、システムは環境に対して開かれたものとみなされてきた。これが、もっとも一般的な意味での開放システムあるいはシステムの開放性である。
ところが、オートポイエーシス理論が登場したとき、提唱者であるマトゥラーナとヴァレラがシステムの閉鎖性を強調したために、混乱や誤解が生じた。二人は生物学者であるから、一般システム理論の提唱者であるベルタランフィが掲げた開放システムとしての生物という見方に真っ向から対立する理論を掲げたかのように思われた(とくに門外漢にとっては)。いったい生物は開放システムなのか閉鎖システムなのか。この初歩的な混乱や誤解は、オートポイエーシスの理論を取り込んだルーマンの社会システム理論の理解にまで混乱や誤解を生み出した。この混乱や誤解を解く鍵は、作動上の閉じという概念にある。この概念の説明をする前に、最初に開放システムと閉鎖システムはどのように考えるべきかという問題を整理しておこう。
まず、論理的に考えて、あるシステムの同一の作動の同一の側面が、環境に対して開いていると同時に閉じているということはありえない、ということを確認しておこう。もしそのようなことがありうるとしたら、そもそも普通の論理学とは異なる前提に立った理論構築が必要になるからである。熱力学的に見ると、完全に閉じたシステムは、エントロピーが最大化して完全な無秩序状態になると考えられている。これが熱力学の第二法則と呼ばれるものであり、エントロピーが最大化したシステムは熱力学的平衡の状態になる。他方、何らかの秩序が成立している状態は、開いたシステムが成立してエントロピー増大を抑制していると考えられ、熱力学的非平衡の状態である。閉じたシステムは、どのような初期状態から出発しても、かならずエントロピーが最大化して熱力学的平衡に達するのに対して、開いたシステムは、すくなくとも一定の時間、秩序を維持し、あるいは秩序を変化させる。
個体としての生物は、十分に長い時間が経てば、かならず死を迎えて土に帰る(つまり完全に無秩序な状態になる)が、生きているあいだは一定の秩序ある状態を保っている。代謝やホメオスタシスはその主要なメカニズムである。そのような生物の生命の本質を、物理学者のE.シュレーディンガーは「生物は負のエントロピーを食べて生きている」と表現した(Schrödinger, 1944=2008)。もう少し正確にいうならば、生物は内部でつねに発生するエントロピーを代謝によって環境に放出し、みずからのエントロピー増大を抑制している、ということになるだろう。そのためには、生物は熱力学的に見て開いたシステムでなければならない。
マトゥラーナとヴァレラも、このことを否定するわけではない。代謝が行われるためには、システムとしての生物有機体と環境とのあいだに物質やエネルギーの交換が成立しなければならず、そのかぎりにおいてシステムは環境に対して開いていなければならない。ベルタランフィが強調したのはこの側面であり、図5では右の入力−出力の部分によって表されている。他方、マトゥラーナとヴァレラが強調したのは、代謝における再生産回路が内部完結しているという側面だった。図5では左の循環的な回路の部分によって表されている。細胞=オートポイエティックな単位体のなかでは、境界づけられたシステムが代謝反応のネットワークを生成し、それが分子構成要素を産出し、それが集まって組織されて境界づけられたシステムを決定し、それが代謝反応のネットワークを生成し、‥‥という過程が、循環的に始まりも終わりもなく続くのである。(Luisi, 2003: p.51)
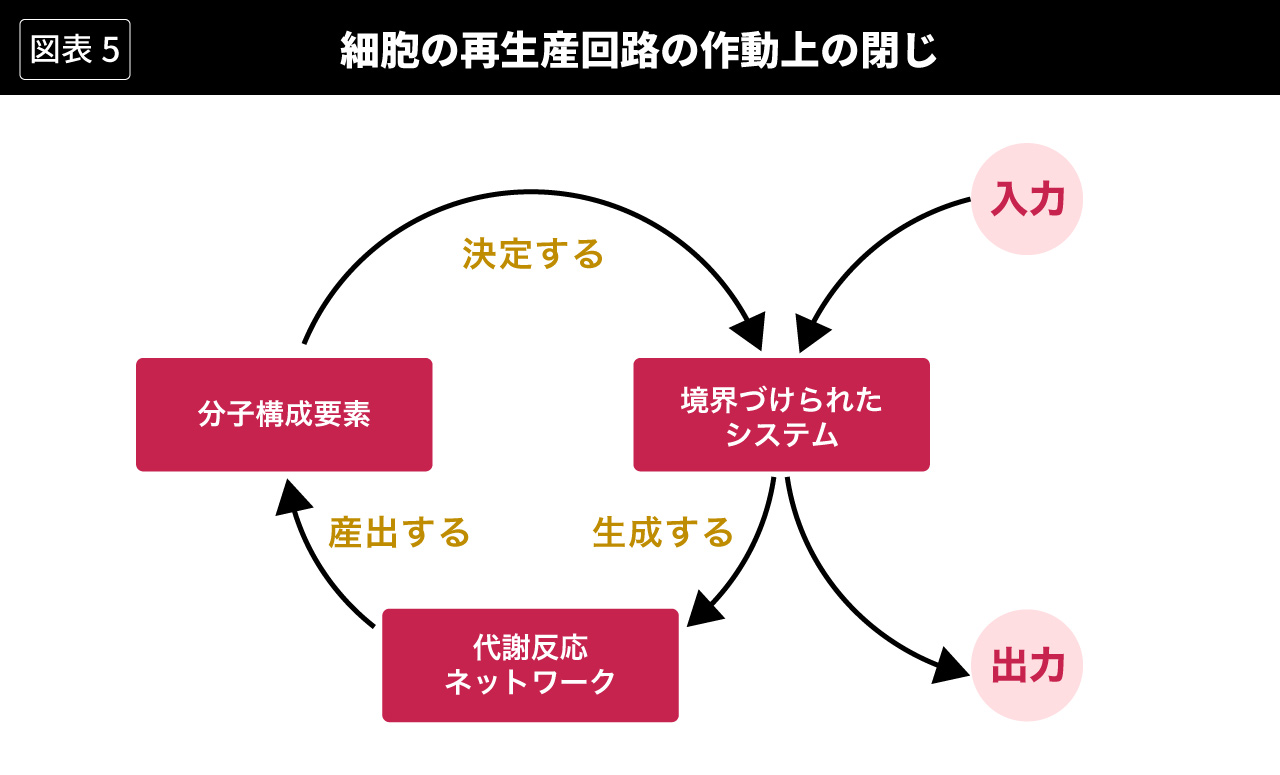
マトゥラーナとヴァレラ自身によるオートポイエーシスの説明は、いきなり読むとかなり難解なのだが、以上のような考え方にもとづけば、比較的理解しやすいだろう。
オートポイエティック・マシンとは、構成素が構成素を産出するという産出(変形および破壊)過程のネットワークとして、有機的に構成(単位体として規定)された機械である。このとき構成素は、次のような特徴をもつ。(ⅰ)変換と相互作用をつうじて、自己を産出するプロセス(関係)のネットワークを、絶えず再生産し実現する、(ⅱ)ネットワーク(機械)を空間に具体的な単位体として構成し、またその空間内において構成素は、ネットワークが実現する位相的領域を特定することによってみずからが存在する。(Maturana/Varela, 1908=1991、訳pp.70-71)
細胞の再生産回路が内部完結している側面を、マトゥラーナとヴァレラは作動上の閉じ(operational closure)と呼んだ。この作動上の閉じという概念が適用されるもう一つの生物の性質が、神経回路の再帰的作動である。生物の進化にともなって、環境の刺激を受けとめる受容器とその刺激に対する反応(行動)を生みだす効果器のあいだに神経回路が発達する。図6の左の状態のように、受容器と効果器がいわば直結している場合には、単純な刺激−反応図式(原因−結果図式)が成立する。しかし、神経回路がしだいに複雑になって図6の右の状態のように受容器と効果器のあいだを媒介するようになると、もはや単純な刺激−反応図式(原因−結果図式)は成立しなくなる。複雑なネットワークを形成する神経回路が内部で再帰的に作動するようになり、作動上の閉じを作りだすからである。(図6は[Foerster, 2002]にもとづく。オートポイエーシス理論にそくしたより詳細な記述としては[Maturana/Varela, 1984=1997、訳pp.164-208]を参照)
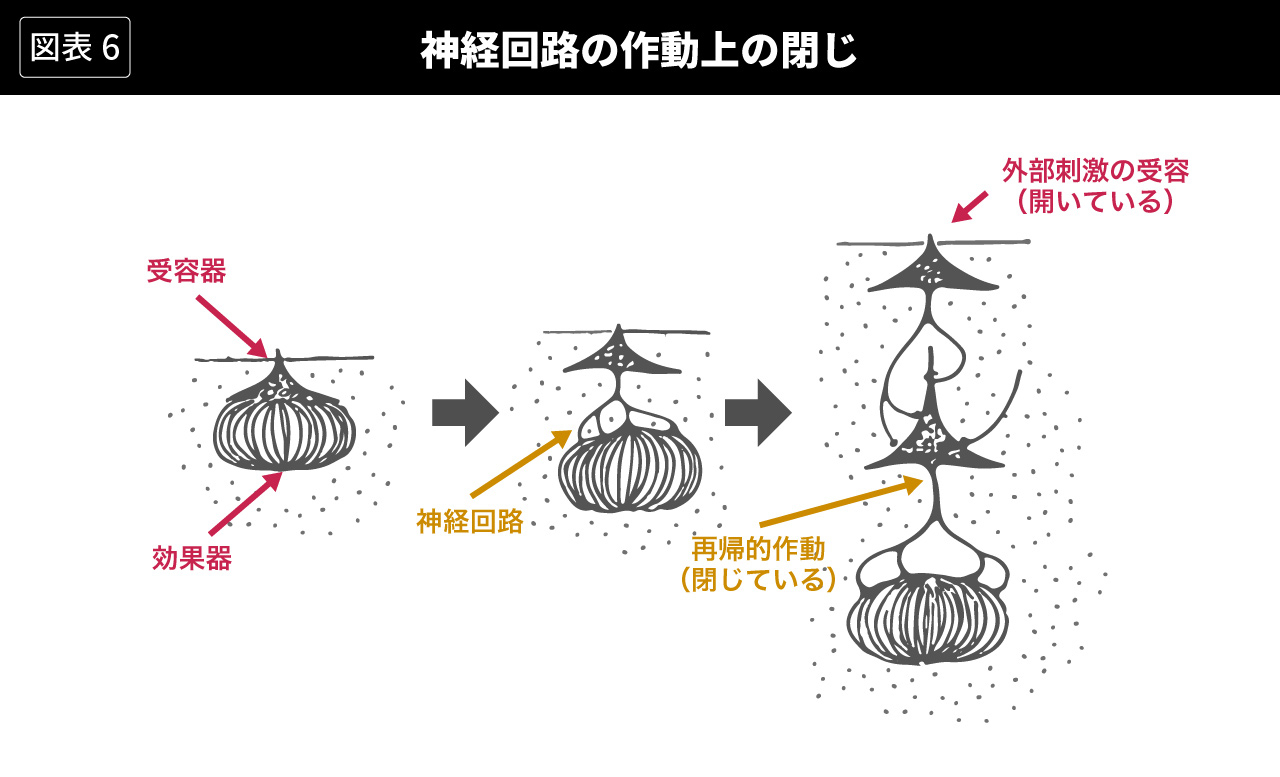
この神経回路の作動上の閉じにもとづいて、マトゥラーナとヴァレラは、生物の認識について、つぎのように要約している。
‥‥神経システムは認識という現象に、ふたつの相互補完的なやりかたで参与する。これらのやりかたは、メタ細胞システムの一部をなしながら作動上の閉じをもつニューロン・ネットワークとしての、神経システムの特別な作動モードにかかわっている。
第一のものは、より明白だ。それは〈有機体がとりうる状態の可能性を拡大すること〉による。この可能性の拡大とは、神経システムが可能にする感覚=運動パターンの大きな多様性──これらまた神経システムの、有機体の作動への参与の鍵をにぎっている──によってもたらされるものだ。
第二のものは、有機体において、有機体がまきこまれているいろいろな[外部との]相互作用と、多くの異なった[有機体の]内部状態とを連合することを可能にする、〈有機体にとって新しい構造的カップリングの次元をひらくこと〉によるものだ。
神経システムの存在あるいは不在が、比較的限定された範囲の認識しかもたない有機体と、人間のように無制限の認識をもつ有機体との、不連続性を決定している。‥‥(Maturana/Varela, 1984=1997、訳pp.206-207:一部改訳)
この記述が、アシュビーとユクスキュルによる最小多様度とシステム/環境−関係についての議論と関連をもつことは、比較的わかりやすいだろう。ユクスキュルの挙げたマダニの例では、神経回路が未発達の単純な生物の場合、みずからの環世界を多様度の小さい単純なものとして認識し、それに対応してみずからの行動の多様度も小さい。それが確実な(種としての)生存戦略を実現している。これに対して、神経回路が発達した人間の場合、みずからの環世界を多様度の大きい複雑なものとして認識し、それに対応してみずからの行動の多様度も大きくすることができる。生物有機体としてのホメオスタシスのメカニズムは、人間であってもとりうる状態の多様度が小さくなるように生物学的に固定されているが、神経システムに媒介される環世界の認識と行動の多様度は、きわめて大きくなりうるはずである。
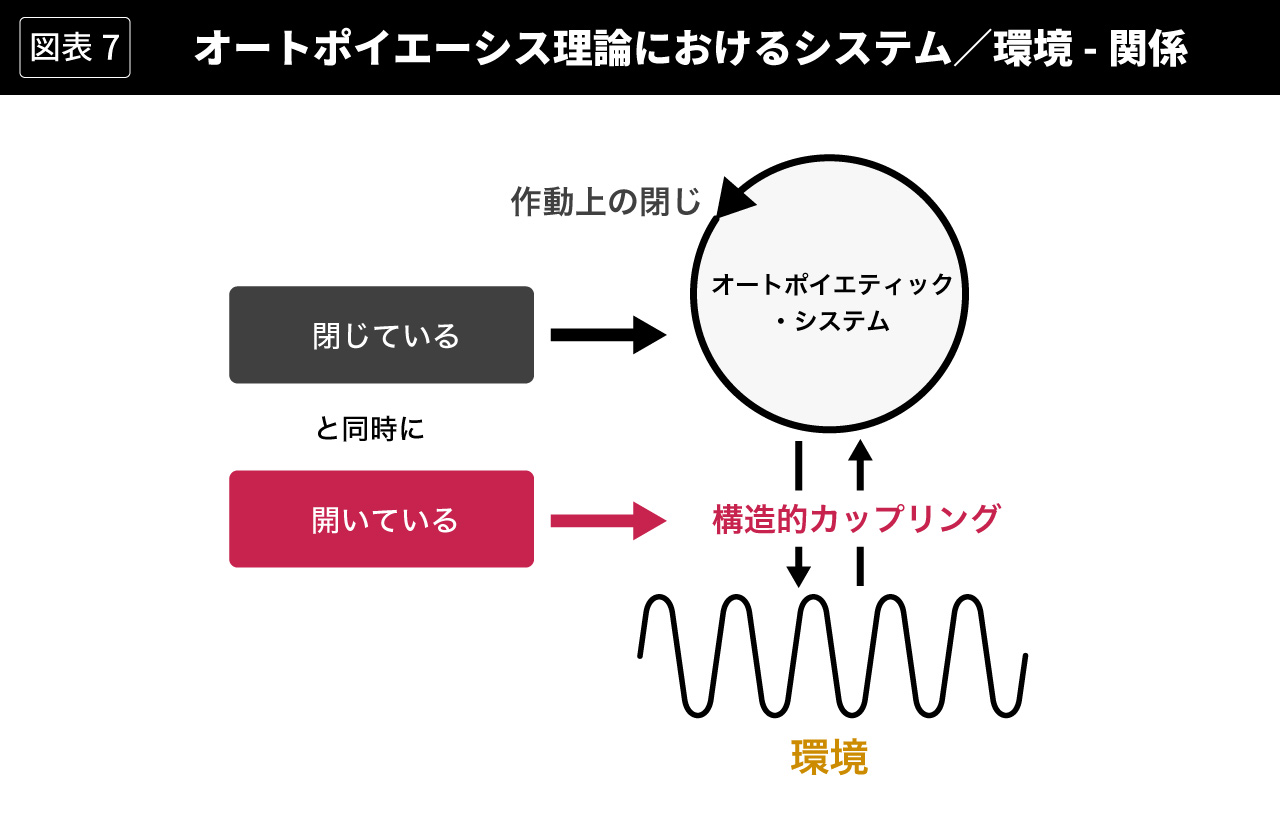
細胞と神経回路についての以上の説明から、オートポイエーシス理論のエッセンスを図式化すると、図7のようになる(図7は[Maturana/Varela, 1984=1997]にもとづく)。オートポイエティック・システムでは、システム内部の回路が再帰的に(循環的に)作動して、閉じた回路を形成している。それが作動上の閉じである。この内部回路は自律的に作動するので、外部環境からの入力によって因果的に規定されない。そのようなシステム/環境−関係を構造的カップリングと呼ぶ。作動上の閉じの側面に注目すれば、システムは環境に対して閉じているということができるし、構造的カップリングの側面に注目すれば、システムは環境に対して開いているということができる。
マトゥラーナとヴァレラは基本的に閉鎖性を強調するが、この理論を社会システム理論に適用したルーマンは、きわめて省略的にシステムは閉じていると同時に開いているという言い方をすることがある。くり返すが、あるシステムの同一の作動の同一の側面が、開放的であると同時に閉鎖的であることは、理論的にありえない。システムのどのような作動のどのような側面に焦点を合わせるかを明確にしないと、システムの開放性/閉鎖性についての議論は混乱や誤解を招くだけである。この点をしっかりと念頭において、次回からはいよいよルーマンの社会システム理論に進むことにしよう。
参考文献
Ashby, Walter Ross, 1956, An Introduction to Cybernetics, Chapman and Hall.(篠崎武・山崎英三・銀林浩 訳『サイバネティクス入門』宇野書店、1967年)
Ashby, Walter Ross, 1960, Design for a Brain: The Origin of Adaptive Behavior, Chapman and Hall.(山田坂仁・宮本敏雄・銀林浩・橋本和美 訳『頭脳への設計』宇野書店、1967年)
Bertalanffy, Ludwig von, 1968, General Systems Theory: Foundations, Development, Applications, George Braziller.(長野敬・太田邦昌 訳『一般システム理論──その基礎・発展・応用』みすず書房、1973年)
Cannon, W. B., 1932, The Wisdom of the Body, W. W. Norton.(舘鄰・舘澄江 訳『からだの知恵』講談社、1981年)
Foerster, Heinz von, 2002, On Constructing a Reality, Understanding Understanding: Essays on Cybernetics and Cognition, Springer.
グロービス経営大学院 MBA用語集、「ポジティブ・フィードバック」(https://mba.globis.ac.jp/about_mba/glossary/detail-11992.html)、「ネガティブ・フィードバック」(https://mba.globis.ac.jp/about_mba/glossary/detail-11900.html)、(2023年4月16日閲覧)
Habermas, Jürgen/Luhmann, Niklas, 1971, Theorie der Gesellschaft oder Sozialtechnologie – Was leistet die Systemforschung?, Suhrkamp.(佐藤嘉一・山口節郎・藤沢賢一郎 訳『批判理論と社会システム理論』木鐸社、1981年)
Luisi, Pier Luigi, 2003, Autopoiesis: a review and a reappraisal, Naturwissenschaften, 90: 49-59.
Maruyama, Magoroh, 1963, The Second Cybernetics: Deviation-Amplifying Mutual Causal Processes, American Scientist 5.2: 164-179.(佐藤敬三 訳『セカンド・サイバネティクス─逸脱増幅相互因果過程』、北川敏男・伊藤重行 編『システム思考の源流と発展』九州大学出版会、1987年: 77-103)
Maturana, Humberto R./Varela, Francisco J., 1980, Autopoiesis and Cognition: The Realization of the Living, D. Reidel.(河本英夫 訳『オートポイエーシス──生命システムとはなにか』国文社、1991年)
Maturana, Humberto R./Varela, Francisco J., 1984, El arbol del conocimiento, Editorial Universitaria.(菅啓次郎 訳『知恵の樹──生きている世界はどのようにして生まれるのか』筑摩書房、1997年)
Parsons, Talcott, 1951, The Social System, Free Press(佐藤勉 訳『社会体系論』青木書店、1974年)
Schrödinger, Erwin, 1944, What is Life?: The Physical Aspect of the Living Cell, Cambridge University Press.(岡小天・鎮目恭夫 訳『生命とは何か──物理的にみた生細胞』岩波書店、2008年)
徳安彰, 2017, 「ホメオスタシス(恒常性維持)」、『社会学理論応用事典』丸善: 466-467
富永健一『思想としての社会学』新曜社、2008年
Uexküll, Jacob von/Kriszat, Georg, 1934, Streifzüge durch die Umwelten von Tieren und Menschen: ein Bilderbuch unsichtbarer Welten, Julius Springer.(日高敏隆・羽田節子 訳『生物から見た世界』岩波書店、2005年)
Wiener, Norbert, 1950, The Human Use of Human Beings: Cybrernetics and Society, Doubleday.(鎮目恭夫・池原止戈夫 訳『人間機械論──人間の人間的な利用』(新装版)みすず書房、2007年)
Wiener, Norbert, 1961, Cybernetics, or, Control and Communication in the Animal and the Machine, MIT Press.(池原止戈夫・彌永昌吉・室賀三郎・戸田巌 訳『サイバネティクス─動物と機械における制御と通信』岩波書店、2011年)


